
Taxonomie du parasite
La position systématique de ce parasite est la suivante :
Ø Embranchement des plathelminthes,
Ø Classe des cestodes,
Ø Sous-classe des Eucéstodes,
Ø Ordre des cyclophyllidés,
Ø Famille des taeniida,
Ø Genre Echinococcus. (Rudolphi, 1801)
Au niveau taxonomique, quatre espèces sont reconnues pour le genre Echinococcus:
– E.granulosus (Batsch , 1786)
– E.multilocularis (Leuckart, 1863)
– E.oligarthrus (Diesing, 1863)
– E.vogeli (Rausch and Bernstein, 1972)
De plus Echinococcus multilocularis présente deux variétés :
– E. multilocularis var.multilocularis, variété européenne décrite par Vogel (1957) : variété » de l’ancien monde »,
– E.multilocularis var.sibiricensis, variété sub-boréale, sibérienne, japonaise et nord-américaine décrite par Rausch et Schiller (1954): variété du « nouveau monde « .
Morphologie du parasite
Le ver adulte dans le tractus digestif de l’hôte définitif
Le stade adulte de E. multilocularis (Figure 1), qui colonise l’intestin grêle de Carnivores, est un cestode de seulement 1,2 à 4,5mm de longueur.
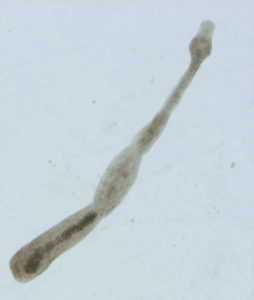
Figure 1 : photographie d’un stade adulte de E.m observé sous loupe binoculaire : x25
Comme tous les cestodes, il ne comporte pas de bouche et tous ses échanges métaboliques se déroulent à travers son tégument.
Dans sa partie antérieure, il possède un organe spécialisé dans la fixation : le scolex, qui porte quatre ventouses musculeuses, une couronne de petits crochets et une de grands crochets, comme c’est visible sur la Figure 2.
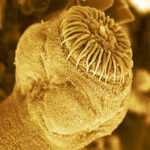
Figure 2 : photographie en microscopie électronique
d’un scolex de E.m.
Le nombre et la taille de ces crochets est un critère de différentiation de E. multilocularis par rapport aux autres espèces du genre Echinococcus (Eckert et al., 2001a).
Ainsi la taille moyenne des grands crochets est comprise entre 24,9 et 34µm, entre 20,4 et 31µm pour les petits (Thompson & McManus, 2001). Ce scolex permet au parasite de se fixer profondément entre les villosités intestinales de l’hôte dans les cryptes de Lieberkühn (Thompson, 1995).
Le strobile est segmenté, formé d’unités appelées proglottis, en nombre compris entre deux et six, très fréquemment cinq (Eckert et al., 2001a).
Les segments portent un pore génital latéral proéminent. La position de ce pore génital est également un critère de diagnose d’espèce. Celui de E. multilocularis est toujours antérieur au milieu du segment (Figure 3).
Le ver adulte est hermaphrodite, et, suite à la fécondation, l’utérus, sacciforme dans cette espèce, se dilate dans le segments terminal par accumulation d’œufs (Thompson & McManus, 2001) (Figure 3).
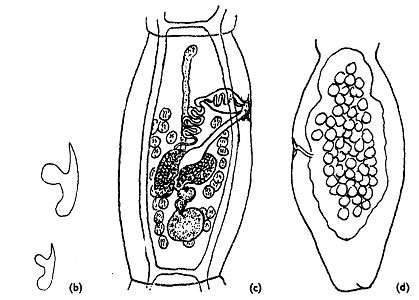
Figure 3 : schéma de l’organisation de portions de E. multilocularis : (b) : crochets (x 700) ; (c) : segment mûr (x 90); (d) : segment ovigère (x 100) ; d’après Euzéby, 1971.
Le site de prédilection de E. multilocularis est la partie postérieure de l’intestin grêle. Cependant, en cas de forte infestation, le parasite peut occuper toutes les portions de cet intestin (Eckert et al.,2001a).
La présence du parasite dans le tube digestif de l’hôte ne cause généralement pas de syndrome particulier. Elle peut provoquer des changements mineurs, tels que l’aplatissement des cellules épithéliales, une légère infiltration cellulaire de la muqueuse et une augmentation de la production de mucus (Eckert et al , 2001a).
On sait globalement peu de choses sur la biologie du parasite chez son hôte principal, probablement à cause du risque que comporte son étude expérimentale. D’après les données les plus récentes, la période prépatente semble être de quatre semaines. Par la suite, l’excrétion d’œufs pourrait durer de un mois et demi à quatre mois (Eckert, 1998, Petavy, 1999). Chaque ver, en période patente, peut produire de 28 à 35 œufs par jour (Thompson & McManus, 2001).
Comme il y a toujours plusieurs vers adultes présents en même temps dans le tube digestif de l’hôte, il y a libération dans les fèces d’un grand nombre d’œufs, libres ou contenus dans les proglottis.
L’œuf dans l’environnement
Lorsqu’ils sont émis dans les fèces, les segments ovigères sont animés de contractions et de relaxations rythmiques qui expulsent les œufs et peuvent les disperser à plusieurs mètres sur le sol ou la végétation environnante (Boucher et al., 2001).
Les œufs du parasite relâchés dans l’environnement par l’hôte principal sont immédiatement infestants. De forme ovoïde (30 à 40 µm de diamètre), ils contiennent un embryon hexacanthe ou oncosphère, premier stade larvaire du parasite. Cet embryon est entouré de plusieurs enveloppes, dont une très kératinisée, appelée embryophore. Celle-ci confère à l’œuf son apparence sombre et striée (Figure 4).
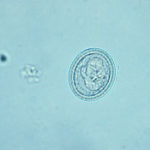
Figure 4 :
aspect microscopique d’un œuf de
E. multilocularis
L’aspect des œufs d’Em ne permet aucune distinction d’avec les œufs de tous les autres membres de la famille des Taeniidae (Thompson & McManus, 2001).
Les œufs de E. multilocularis sont dotés d’une forte capacité de résistance dans le milieu extérieur. Même après plusieurs semaines, ils demeurent infestants lorsqu’un hôte intermédiaire potentiel les consomme.
La larve dans le parenchyme hépatique de l’hôte intermédiaire
L’hôte intermédiaire, généralement un Rongeur ou un Insectivore, s’infeste en ingérant des œufs.
Suite à l’action des enzymes de l’estomac et de l’intestin grêle, l’oncosphère est libérée de son embryophore (Thompson, 1995). La bile joue un rôle en activant l’oncosphère qui pénètre dans la paroi de l’intestin grêle, à l’aide des mouvements de ses crochets et de possibles sécrétions. Après avoir gagné une veinule, l’oncosphère est transportée passivement jusqu’au foie de l’hôte. Le métacestode s’y développe primairement (Thompson & Mc Manus, 2001).
Ce deuxième stade larvaire consiste en un agrégat de petites vésicules, de un à dix millimètres de diamètre en moyenne (parfois jusqu’à 20 ou même 30mm) qui forment une structure alvéolaire (Deplazes & Eckert, 2001). L’aspect extérieur de cette larve est visible sur la Figure 5.
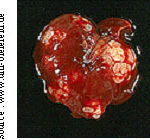
Figure 5 : foie de M. arvalis
parasité par E.m.
Ce métacestode a la capacité de proliférer par bourgeonnement externe, formant des ramifications qui permettent au parasite d’envahir progressivement les organes de l’hôte (Thomson, 1995 ; Eckert, 1998).
Ce tissu parasitaire invasif semble être protégé des réactions immunitaires de l’individu par la membrane basale, riche en hydrates de carbone, et contenant un antigène nommé antigène Em2 (Gottstein et al., 1992).
Le détachement de cellules germinales à partir des protrusions de tissu parasitaire, et leur distribution par les vaisseaux sanguins ou lymphatiques permettent la formation de foyers métastatiques à distance, caractéristiques de E. multilocularis (Amman & Eckert, 1995 ; Eckert, 1998).
Chez les hôtes intermédiaires naturels, les vésicules comportent une membrane germinative qui produit par multiplication asexuée de nombreux protoscolex (Figure 6), forme infestante pour les Carnivores.
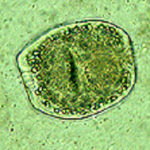
Figure 6 :
protoscolex de E.m. en microscopie optique
C’est donc par la consommation d’hôtes intermédiaires contaminés (Rongeurs, Insectivores) que les Carnivores contractent le parasite. Le développement du métacestode est rapide chez ce type d’hôte intermédiaire et les premiers protoscolex peuvent être produits en 40 à 45 jours (Eckert, 1998). Il s’agit probablement là d’une adaptation à la courte durée de vie de l’hôte (Eckert et al., 2001a). La mort de l’hôte survient généralement au bout de cinq mois .
Le nombre de protoscolex par lésion est très variable, on a montré par exemple des valeurs allant de 14 à 244400 chez Arvicola terrestris (Stieger et al., 2002). Chaque protoscolex est susceptible de se développer en un ver adulte dans l’intestin d’un hôte favorable.
Lorsque des protoscolex sont ingérés par un hôte définitif potentiel, après avoir subit l’action des pepsines de l’estomac, ils se dévaginent dans la partie proximale du duodénum, sous l’effet du changement de pH, de l’exposition à la bile et de l’élévation de la température. Ils se fixent alors à la paroi du tube digestif et se développent en vers adultes prêts à se reproduire (Thompson & McManus, 2001).
Chez les hôtes atypiques (dont l’Homme), la larve est le plus souvent stérile. Il n’y pas de formation de protoscolex et on observe des zones de calcification et de nécrose (Deplazes & Eckert, 2001).